
Detailed Description
 The NaKATPase or Sodium Pump uses one ATP to perform the electrogenic exchange of 2 potassium ions from outside to inside the cell in exchange for 3 sodium ions from inside to out. The pump is reversible, so that ATP can be produced from ADP and Pi when the ionic gradients are suitable and there is adequate ADP available. There are many possible configurations for a model of the pump, depending on the numbers of steps in the binding of Na or K and the places in the sequence of forms for the binding and release of ATP, ADP and Pi. This model by Chapman et al (1983) is a minimal model in the sense that it lumps to the binding of 3 Na into one step, and the binding of 2 K into one step. See the diagram.
The NaKATPase or Sodium Pump uses one ATP to perform the electrogenic exchange of 2 potassium ions from outside to inside the cell in exchange for 3 sodium ions from inside to out. The pump is reversible, so that ATP can be produced from ADP and Pi when the ionic gradients are suitable and there is adequate ADP available. There are many possible configurations for a model of the pump, depending on the numbers of steps in the binding of Na or K and the places in the sequence of forms for the binding and release of ATP, ADP and Pi. This model by Chapman et al (1983) is a minimal model in the sense that it lumps to the binding of 3 Na into one step, and the binding of 2 K into one step. See the diagram.
This model was rewritten by JB Bassingthwaighte (2002) to add:
1. Cell of volume Vi surrounded by medium of volume Vo.
2. Transporter mass constrained to fixed Etot.
3. Mass Balance check for Na, K, and total P in all forms (organic or inorganic)
4. In this version, transporter is on membrane.
5. ATP, ADP, and Piin are incorporated.
6. Added EGoldman and EpredSS for comparison with Vm.
Note: PI (all in caps) is the irrational number, 3.1415926… . Pi is the chemical species, inorganic phosphate.
Parameter Sets
There are many different parameter sets saved within this JSim project file, modeling different conditions for a Sodium Potassium pump. The provided plots from each parameter set is displayed below, and the corresponding model is accessible through JSim (e.g. Figure 2A can be acquired by running the .proj file 2A). Models numbered 5 and greater have the corresponding parameter sets saved within the .proj file (e.g. Figure 5A can be acquired by running the .proj file with parameter set 5A).
Note: Run loops in JSim to acquire the figures displayed below.
Figure 2
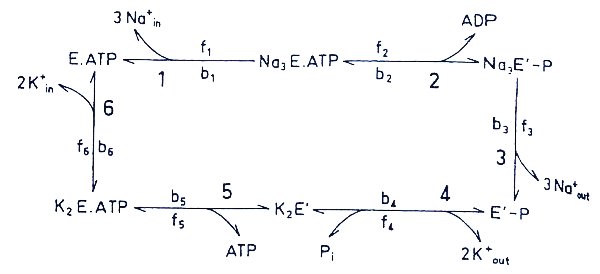
Soup kinetics: examining turnover of ATP by the enzyme in a broth so that the concentrations of Na and K are the same on the release and binding steps. This is a "demand" pump, requiring both substrates to operate. The bottom panel shows ATP flux over the range of concentrations when the sum [Na] + [K] = 150 mM.
-
- 2A: Nain - Naout; Ko - Ki - fixed, varied
-
- 2B: Ko - Ki; Nain - Naout varied
-
- 2C: Nain - Naout; Ko - Ki - 150-Nain
Figure 3
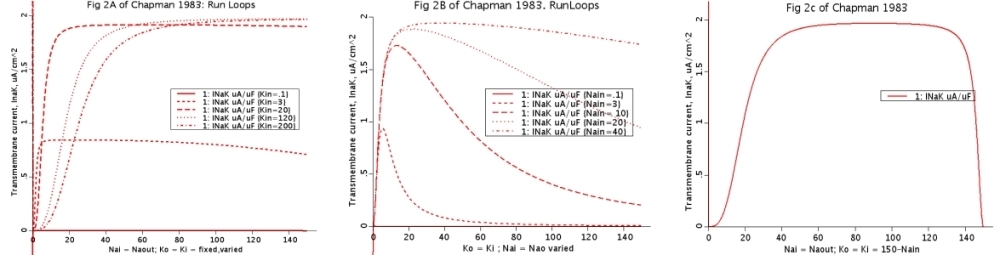
Being an integral membrane protein that transports a charged species, the pump transporter molecule is subject to the influences of transmembrane voltage. Three arrangements are explored in Figure 3, two placing the influence solely at one or other of two particular steps (step 3 or 5) in the sequence. The third puts the influence shared between two steps. Data acquired experimentally suggests that Step 5 is the best choice.
Note: Figure 3C does not match the curve given in the Chapman paper. When the model is run according to the parameters provided by Chapman et al, the curve displayed on this page results. The curve in the Chapman paper has not been reproducible.
-
- 3A: Voltage dependence on step 3
-
- 3B: Voltage dependence on step 5
-
- 3C: Voltage dependence on steps 3 and 5
Figure 4
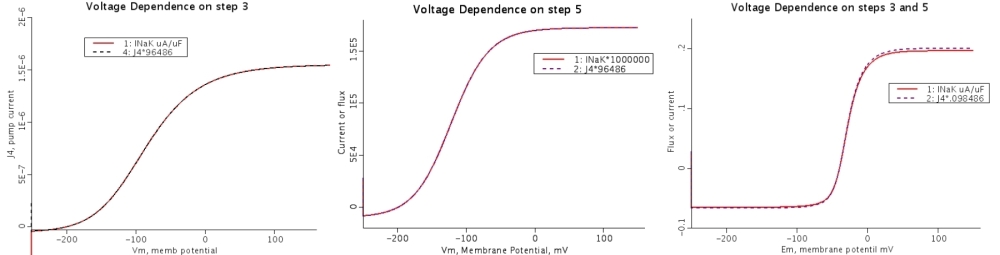
Pump reversibility. In steady state the fluxes through all of the steps are the same. The enzyme recycles without loss. At the pump reversal potential, in the case of the "standard" concentrations for the model, this occurs at -200mV. Changes in parameter values changes the slope of the relationship, not the position of the equilibrium. As can be seen in 4B, less physiological slope conductance is associated with a relatively high degree of voltage-dependent reversal. Conversely, 4C shows that a larger physiological slope conductance is associated with little voltage-dependent reversal (Chapman et al, 145).
-
- 4A: Control, with conditions as in Fig. 3B
-
- 4B: Rate coefficients adjusted to increase the voltage-dependent reversibility exhibited by curve A. b5 decreased by a factor of 30.
-
- 4C: Rate coefficients adjusted to reduce the reversibility. f5 decreased by a factor of 10, f6 decreased by a factor of 100, b6 decreased by a factor of 104.
Figure 5
Looking for constraints from data. Parametric configurations B and C in Figure 4 are both unreasonable, as judged by “soup kitchen kinetics”. (Non-vectorial means no direction, in the soup.)
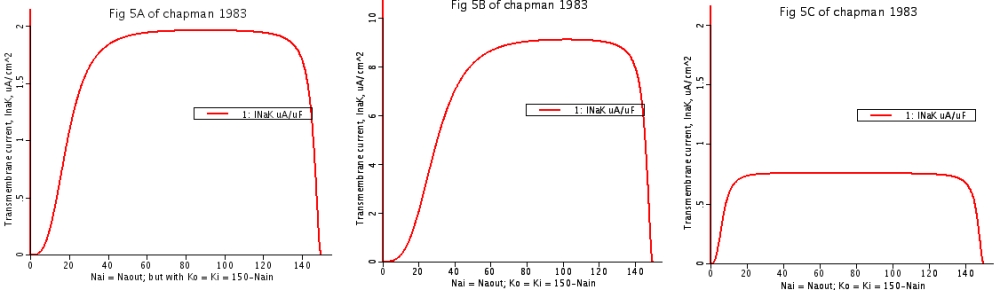
-
- 5: Simultaneous activation of non vectorial ATPase activity by Na+ and K+ ions constrained to sum to 150 mM, using rate coefficients yielding the current-voltage relations of Figure 4A, B, and C.
Figure 6
Energetics. The equilibrium potential is shifted almost +100 mV by reducing ATP by 40% and raising ADP to maintain total nucleotide level. Effects of altering the chemical conditions. Relative irreversibility totally disappears with reduction in ATP concentration from 6A to 6B. 6B shows that voltage is a powerful force in the synthesis of ATP, under conditions of reduced initial ATP concentrations. As can be seen from 6C, an increase of ionic concentration gradients does not produce more significant pump reversal than imposition of a voltage gradient. The forward rate of reaction is simply reduced.
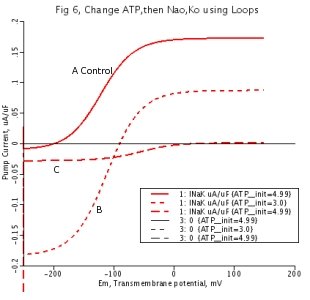
6A: Control, [ATP] = 4.99 mM
6B: Reducing [ATP] from 4.99 mM to 3.0 mM, corresponding increase in [ADP]
6C: Increasing ionic concentration gradients. [Na+] increased to 200mM, extracellular [K+] decreased to 0.1mM
Figure 8
Interrelationships between ionic concentrations and ATP levels. Analyzing the relationship between ATP concentrations and the current experienced across the membrane, via the transport of K+out. All rate coefficients not explicitly mentioned are as in the .proj file 3B with [ATP] + [ADP] constrained to sum to 5.05mM.
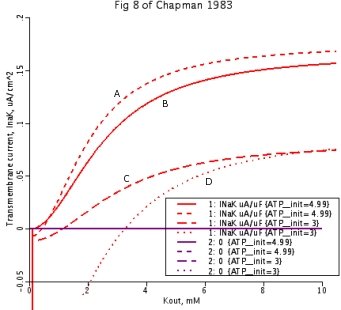
8A: [ATP] set at 4.99 mM, [Na+]out set at 50 mM.
8B: [ATP] set at 4.99 mM, [Na+]out set at 140 mM.
8C: [ATP] set at 3.0 mM, [Na+]out set at 50 mM.
8D: [ATP] set at 3.0 mM, [Na+]out set at 140 mM.
Relevant Equations
The following complete equation is derived from annotated equations in the JSim model.
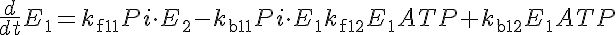
JSim Tutorial
Click here to go to a JSim tutorial webpage, with an introduction to the JSim GUI, detailed usage instructions, and an accompaying video.
J. Brian Chapman, Edward A Johnson, and J. Mailen Kootsey, Electrical and Biochemical Properties of an Enzyme Model of the Sodium Pump, Journal of Membrane Biology 74., 139-153, 1983
P.Lauger. Electrogenic Ion Pumps, Sinauer, Sunderland MA 1991 pp313
WD Stein. Channels Carriers and Pumps, Academic Press 1990 pp325
RM Gulrajani. Bioelectricity and Biomagnetism, Wiley, New York pp729
N. Sperelakis. Origin of the cardiac resting potential. Handbook of Physiology, Sec. 2, The Cardiovascular System. Vol. 1,, Waverly Press, Inc., 1979 pp187-267
Copyright (C) 1999-2009 University of Washington. From the National Simulation Resource, Director J. B. Bassingthwaighte, Department of Bioengineering, University of Washington, Seattle WA 98195-5061. Academic use is unrestricted. Software may be copied so long as this copyright notice is included. This software was developed with support from NIH grant HL073598. Please cite this grant in any publication for which this software is used and send one reprint to the address given above.
Model development and archiving support at https://www.imagwiki.nibib.nih.gov/physiome provided by the following grants: NIH U01HL122199 Analyzing the Cardiac Power Grid, 09/15/2015 - 05/31/2020, NIH/NIBIB BE08407 Software Integration, JSim and SBW 6/1/09-5/31/13; NIH/NHLBI T15 HL88516-01 Modeling for Heart, Lung and Blood: From Cell to Organ, 4/1/07-3/31/11; NSF BES-0506477 Adaptive Multi-Scale Model Simulation, 8/15/05-7/31/08; NIH/NHLBI R01 HL073598 Core 3: 3D Imaging and Computer Modeling of the Respiratory Tract, 9/1/04-8/31/09; as well as prior support from NIH/NCRR P41 RR01243 Simulation Resource in Circulatory Mass Transport and Exchange, 12/1/1980-11/30/01 and NIH/NIBIB R01 EB001973 JSim: A Simulation Analysis Platform, 3/1/02-2/28/07.