
One Enzyme 2 Substrates, A and B, to 2 Products, C and D, Reversible Second-order enzyme kinetic model with capacitance and reversible enzymatic reaction with binding and release kinetics for two substrates givng two products, thermodynamically constrained.
Description
This model represents the enzymatic conversion of two substrate, A and B, to two products, C and D.
First is the binding of one substrate to the enzyme, E, to form EA or EB, a 1-substrate-enzyme complex, ES.
This binding is followed by binding the other substrate to form EAB. This is random bi-bi, not ordered.
Next is a reaction-release event, which yields one product, C or D, and an enzyme still bound to the
other product. The second release yields the free enzyme and the second product.
Note 1. The equilibrium between EAB and E is independent of the route taken: the alp1 and alp2 factors
are to assure this. Because there is no disinction between EAB and ECD in this model alp2 = alp1 in order to
keep the same ratio EAB/E = ECD/E.
Note 2. The model parameters can be adjusted to fit Michaelis-Menten conditions (Substrate
concentrations high compared to enzyme; on/off reaction fast compared to forward
reaction rate) and Briggs-Haldane conditions where the On-rate is low. Reducing the [Enzyme] relative
to the substrate and product concnetrations reduces the effect of the capacitance of the enzyme-bound
forms on the kinetics of the reactions. This results in faster attainment of the steady state.
Note 3: Consuming the products to reduce the back flux drains the system; Segel puts all products to zero
in order to calculat the M-M equivalent equations.
When without a drain, the system --> equilibrium, as in this version. EquilA is the theoretical value.
Note 4. Substrate is preserved: Stot: Sum at beginning = Sum at end.
Enzyme is conserved: Etest.
Note 5. This code converts the model from Random Bi-Bi to Ordered Bi Bi
simply by putting an intital binding, k1 or j1 to zero, and likewise
putting K_4 or j_4 to zero.
Note 6. Segel's approach is instantaneous steady state, i.e. the binding and release
rates are exceeding fast, so his reaction velocities, shown by v and by v/Vmax,
are reached instantaneously even as concentrations change. Our approach here is to
account for the binding and unbinding that actaully take time, and alos to account
for the capacitance in the enzymatic reactions by accounting for the substrates an
products bound to the enzyme. This is most important with high enyme concentrations
or low substrate concnetrations in the range of the binding affinities.
Note 7. Observe that EAB achieves a peak concentration at early times when A and B
are first sliding through the series of reaction steps starting from A and B with C
and D = 0. This peak is always(?) higher than at equilibrium.
Simultaneous substrate binding and simultaneous product release bidirectional:
Net reaction A + B <----> C + D E = Enzyme concentration
k1 k2 k3 k4
A+E <--> EA + B <-->\ /<----> EC + D <---> E + C
k_1 k_2 \ / k_3 k_4
EAB=ECD
j1 j2 / \ j3 j4
B+E <--> EB + A <-->/ \<----> ED + C <---> E + D
j_1 j_2 j_3 j_4
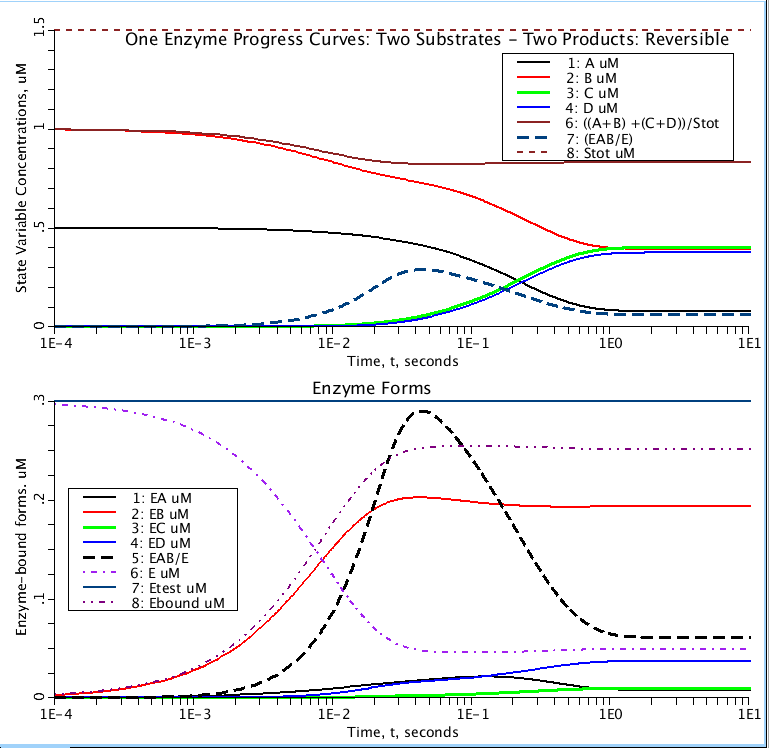
Figure: Progress curves of substrates (A,B), products (C,D), and enzyme (E). Etest is Ebound plus E (unbound). Stot is total amount of substrates and products.
Equations
The equations for this model may be viewed by running the JSim model applet and clicking on the Source tab at the bottom left of JSim's Run Time graphical user interface. The equations are written in JSim's Mathematical Modeling Language (MML). See the Introduction to MML and the MML Reference Manual. Additional documentation for MML can be found by using the search option at the Physiome home page.
- Download JSim model MML code (text):
- Download translated SBML version of model (if available):
We welcome comments and feedback for this model. Please use the button below to send comments:
These references do not include either the capacitance or the kinetics of binding or release.
Segel, I. H. Biochemical Calculations. 2nd Ed.Wiley 1976, p.294.
Segel, I. H. Enzyme Kinetics> Behavior and Analysis of RApid Equilibria and Steady-State Enyme Systems
Wiley 1993 p 274-319
Bassingthwaighte JB.: Enzymes and Metabolic Reactions, Chapter 10 in "Transport and Reactions
in Biological Systems", Pages 7-8
Please cite https://www.imagwiki.nibib.nih.gov/physiome in any publication for which this software is used and send one reprint to the address given below:
The National Simulation Resource, Director J. B. Bassingthwaighte, Department of Bioengineering, University of Washington, Seattle WA 98195-5061.
Model development and archiving support at https://www.imagwiki.nibib.nih.gov/physiome provided by the following grants: NIH U01HL122199 Analyzing the Cardiac Power Grid, 09/15/2015 - 05/31/2020, NIH/NIBIB BE08407 Software Integration, JSim and SBW 6/1/09-5/31/13; NIH/NHLBI T15 HL88516-01 Modeling for Heart, Lung and Blood: From Cell to Organ, 4/1/07-3/31/11; NSF BES-0506477 Adaptive Multi-Scale Model Simulation, 8/15/05-7/31/08; NIH/NHLBI R01 HL073598 Core 3: 3D Imaging and Computer Modeling of the Respiratory Tract, 9/1/04-8/31/09; as well as prior support from NIH/NCRR P41 RR01243 Simulation Resource in Circulatory Mass Transport and Exchange, 12/1/1980-11/30/01 and NIH/NIBIB R01 EB001973 JSim: A Simulation Analysis Platform, 3/1/02-2/28/07.